
To differentiate between early and late-stage apoptotic events, simultaneous staining with fluorescently-labeled Annexin V and viability dyes such as 7-Aminoactinomycin D (7-AAD) or propidium iodide (PI) can be used. Annexin V (molecular weight of 35-36 kDa) is a human vascular anticoagulant that is a Ca2+ dependent phospholipid-binding protein with a high affinity for the anionic phospholipid phosphatidylserine (PS). In healthy cells, PS is localized to the cytosolic side of the plasma membrane, but during the early stages of apoptosis, it is translocated to the extracellular side. This process makes Annexin V staining useful for detecting the early stages of apoptosis.
Viability dyes such as 7-AAD and PI rely on the loss of membrane integrity for entry into the cell and will not stain cells during early apoptotic events when membranes are still intact. Therefore, cells in the early stages of apoptosis can be identified as positive for Annexin V staining and negative for 7-AAD or PI staining. In contrast, cells in the later stages of apoptosis will be characterized by Annexin V staining along with staining with 7-AAD or PI. This simultaneous staining approach provides a sensitive and reliable method for distinguishing between early and late-stage apoptotic events and can be useful in a wide range of applications, including drug discovery and monitoring of cancer treatment efficacy.
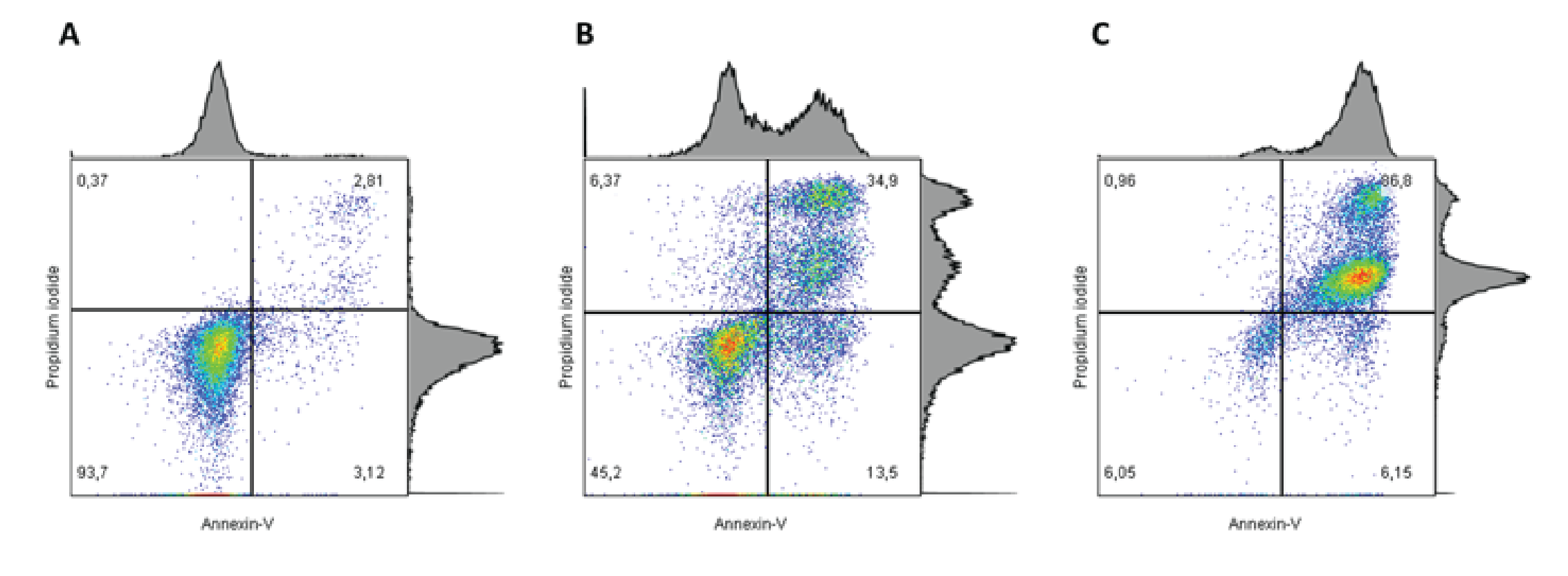
Figure 1: Cells were stained with APC-conjugated Annexin V and PI. The proportion of apoptotic events increases with the concentration of drug treatment i.e. from A→C. In addition, a higher proportion of early stage apoptosis is observed with an intermediate concentration of drug treatment (B) relative to the highest concentration (C). The most late-stage apoptotic events are observed with the highest concentration of drug treatment (C).
REFERENCES:
Ranganathan, P., Jayakumar, C., Navankasattusas, S., Li, D. Y., Kim, I., & Ramesh, G. (2014). UNC5B receptor deletion exacerbates tissue injury in response to AKI. Journal of the American Society of Nephrology, 25(2), 239–249. https://doi.org/10.1681/ASN.2013040418
Wang, Y., Zhao, J., Yang, W., Bi, Y., Chi, J., Tian, J., & Li, W. (2015). High-dose alcohol induces reactive oxygen species-mediated apoptosis via PKC-β/p66Shc in mouse primary cardiomyocytes. Biochemical and Biophysical Research Communications, 456(2), 656–661. https://doi.org/10.1016/j.bbrc.2014.12.012
Yu, J., Zheng, J., Zhao, X., et al. (2014). Aminoglycoside stress together with the 12S rRNA 1494C>T mutation leads to mitophagy. Lightowlers, R. (Ed.). PLoS ONE, 9(12), e114650. https://doi.org/10.1371/journal.pone.0114650
Caspases are a family of cysteine-aspartic proteases that play a crucial role in the execution of apoptosis. Caspase activation can be assessed using flow cytometry through various methods, including indirect measures, such as DEVD peptides and identification of cleaved Poly (ADP-ribose) polymerase (PARP) and the fluorescent inhibitor of caspases (FLICA) methodology. The amino acid sequence DEVD (Aso-Gly-Val-Asp) corresponds to a sequence within the PARP enzyme that is cleaved by caspase-3, -6, and -7. DEVD sequences conjugated to specific molecules (DEVD-CHO) may be used to inhibit caspase activity and as indicators of caspase activity when cleaved. In another example, when DEVD is bound to a nucleic acid binding dye, the cleavage of this conjugate releases the nucleic acid binding dye, and the subsequent fluorescent DNA signal is measured using flow cytometry. FLICA reagents include a caspase inhibitor that covalently and irreversibly binds to the reactive cysteines of active caspases. This caspase inhibitor is linked to a fluorescent dye, making it possible to identify the relative extent of caspase activation via the intensity of the fluorescent signal detected by flow cytometry.
REFERENCES:
Kaufmann, S. H., Karp, J. E., Svingen, P. A., Krajewski, S., Burke, P. J., Gore, S. D., & Reed, J. C. (2008). Apoptosis-associated caspase activation assays. Methods, 44(3), 262–272. https://doi.org/10.1016/j.ymeth.2007.11.005
Köhler, C., Orrenius, S., & Zhivotovsky, B. (2002). Evaluation of caspase activity in apoptotic cells. Journal of Immunological Methods, 265(1-2), 97–110. https://doi.org/10.1016/S0022-1759(02)00073-X
Martin, S. J. (2008). Getting the measure of apoptosis. Methods, 44(3), 197–199. https://doi.org/10.1016/j.ymeth.2008.02.001
Another hallmark of apoptosis is the fragmentation of genomic DNA. To monitor DNA breakdown, fluorescent molecule-conjugated deoxyuridine triphosphate nucleotides (dUTPs) can be used to label the 3′-hydroxyl termini of the resulting DNA fragments. The enzyme terminal deoxynucleotidyl transferase (TdT) is added to the assay to catalyze the addition of these dUTPs to the 3′-hydroxyl ends of the DNA fragments. During the analysis, an increase in DNA fragmentation will result in the detection of an increased fluorescent signal. This assay is referred to as the terminal deoxynucleotidyl transferase dUTP nick end labeling assay or TUNEL assay.
REFERENCE:
Darzynkiewicz, Z., Galkowski, D., & Zhao, H. (2008). Analysis of apoptosis by cytometry using TUNEL assay. Methods, 44(3), 250–254. https://doi: 10.1016/j.ymeth.2007.11.008
The loss of mitochondrial function is a major apoptotic event characterized by a loss of the potential charge across the membrane and can be assessed using JC-1, a cationic carbocyanine dye. In healthy cells, JC-1 aggregates within the mitochondria, emitting red fluorescence at approximately 590 nm. When the mitochondria depolarize, JC-1 aggregation diminishes, resulting in monomeric JC-1 accumulation in the cytoplasm and green fluorescence at 529 nm. This metachromatic shift from red to green fluorescence can be easily measured by flow cytometry with blue laser excitation at 488 nm.
REFERENCES:
Liu, T., Singh, R. K., Maity, T. K., Seo, E. Y., & Tavakoli-Yaraki, M. (2007). Flex-Hets differentially induce apoptosis in cancer over normal cells by directly targeting mitochondria. Molecular Cancer Therapeutics, 6(6), 1814–1822. https://doi.org/10.1158/1535-7163.MCT-06-0655
Proost, I., Robertson, H., Stanton, P. K., & Wu, J. (2008). Functional live cell imaging of the pulmonary neuroepithelial body microenvironment. American Journal of Respiratory Cell and Molecular Biology, 39(2), 180–189. https://doi.org/10.1165/rcmb.2007-0316OC
St. John, J. C., Ramalho-Santos, J., Gray, H. L., & Petrosko, P. (2006). The analysis of mitochondria and mitochondrial DNA in human embryonic stem cells. Methods in Molecular Biology, 331, 347–374. https://doi.org/10.1385/1-59745-027-0:347
The cleavage of poly ADP-ribose polymerase (PARP), which is a ubiquitous DNA repair enzyme, is another important characteristic of apoptosis. Caspases-3, -6, and -7 cleave PARP from its 116 kDa form to two fragments measuring 85 kDa and 25 kDa. The cleaved form of PARP can be identified through flow cytometry by using fluorescently-tagged antibodies directed specifically against the cleaved fragments of PARP.
REFERENCE:
Nicholson, D. D. W., Ali, A., Thornberry, N. A., Vaillancourt, J. P., Ding, C. K., Gallant, M., Gareau, Y., Griffin, P. R., Labelle, M., Lazebnik, Y. A., Munday, N. A., Raju, S. M., Smulson, M. E., Yamin, T. T., Yu, V. L., Miller, D. K., & Fesik, S. W. (2002). Identification and inhibition of the ICE/CED-3 protease necessary for mammalian apoptosis. Nature, 376(6535), 37–43. https://doi.org/10.1038/376037a0
Wlodkowic, D., Skommer, J., & Darzynkiewicz, Z. (2009). Flow cytometry-based apoptosis detection. Methods in molecular biology (Clifton, NJ), 559, 10.1007/978-1-60327-017-5_2. https://doi.org/10.1007/978-1-60327-017-5_2
Wlodkowic, D., Telford, W., Skommer, J., & Darzynkiewicz, Z. (2011). Apoptosis and Beyond: Cytometry in Studies of Programmed Cell Death. Methods in Cell Biology, 103, 55–98. https://doi.org/10.1016/B978-0-12-385493-3.00004-8
BD Biosciences. (n.d.). Apoptosis Applications Brochure. Retrieved from https://www.bdbiosciences.com/documents/BD_Apoptosis_Brochure.pdf
De Novo Software. (n.d.). Flow Cytometry Text Book. Retrieved from http://flowbook.denovosoftware.com/
eBioscience. (n.d.). Apoptosis Research Product Guide. Retrieved from https://www.thermofisher.com/us/en/home/life-science/cell-analysis/apoptosis-research-products-guide.html
Fisher Scientific. (n.d.). Molecular Probes Apoptosis/caspase Assays for Flow Cytometry. Retrieved from https://www.thermofisher.com/us/en/home/life-science/cell-analysis/flow-cytometry/flow-cytometry-learning-center/flow-cytometry-resource-library/protocols/molecular-probes-apoptosis-caspase-assays-for-flow-cytometry.html

